Konjugation (Biologie)
- Wikipedia-de: Konjugation (Biologie)
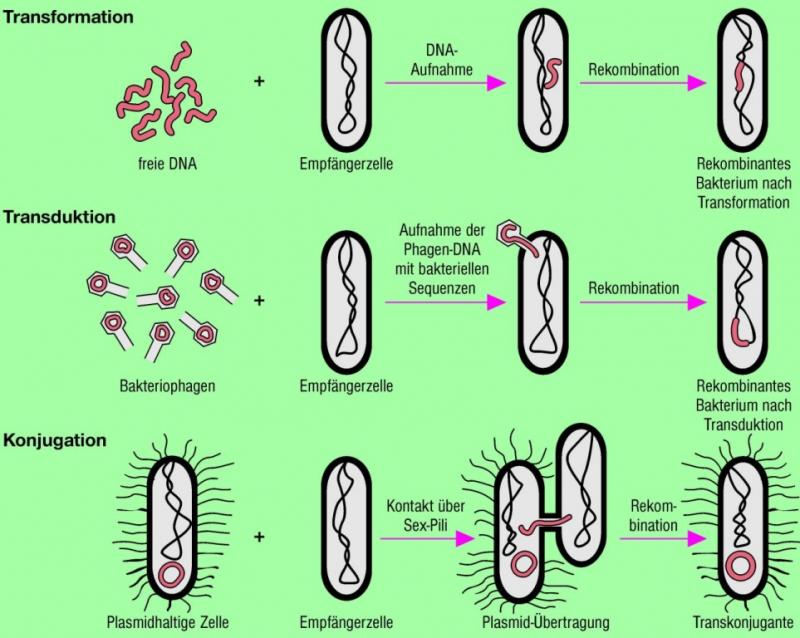
Konjugation (lateinisch coniugare paarweise zusammenbinden) bezeichnet in der Mikrobiologie die Übertragung von Teilen des Genoms von einer Spenderzelle (Donor) auf eine Empfängerzelle (Rezipient) durch direkten Zellkontakt. Darüber hinaus wird die Bezeichnung in der Mykologie, der Lehre von den Pilzen, für die geschlechtliche Verschmelzung spezialisierter Pilzhyphen verwendet.
Konjugation bei Prokaryoten
Die Konjugation bei Prokaryoten ist ein Beispiel für Parasexualität und ist bei ihnen auch weit über die Artgrenzen hinweg möglich. Dabei können beispielsweise Gene für Antibiotikaresistenzen, Toxine und Kolonisierungsfaktoren von einer für den Menschen harmlosen Spezies auf eine pathogene Spezies übertragen werden.
Auch Gene für die Ausbildung von Wurzelknöllchen und Bakteriozine werden durch Konjugation übertragen.
Es gibt außerdem Fälle, in denen DNA von Prokaryoten auf Eukaryoten übertragen wird, z. B. überträgt das Bakterium Agrobacterium tumefaciens sog. Ti-Plasmide in verwundete Stellen bestimmter Pflanzen. In Experimenten wurden auch Konjugationen von Escherichia coli auf die Bäckerhefe, Saccharomyces cerevisiae und auf Eizellen von Hamstern nachgewiesen.
Entdeckt wurde der Vorgang der Konjugation 1946 von Joshua Lederberg und Edward Tatum bei Escherichia coli.
Gramnegative Bakterien
Die Konjugation bei gramnegativen Bakterien wird von Plasmiden, meist in sich geschlossenen DNA-Molekülen mit nicht essentiellen Genen, gesteuert.
Konjugative Plasmide tragen die spezifischen Informationen der Konjugationsmechanismen, d. h. die dafür codierenden Gene in der tra-Region (tra für Transfer), darunter eine Relaxase zum einzelsträngigen Schnitt der zu übertragenden DNA.
Nicht alle Plasmide haben eine tra-Region, es gibt also auch nicht konjugative Plasmide.
Eine Donorzelle ist eine Zelle mit einem konjugativen Plasmid, sie kann mit einem F-Pilus, einem fadenförmigen Zellanhängsel, welches ebenfalls durch Gene auf dem Plasmid codiert wird, Kontakt zu einer Zelle aufnehmen, die dieses Plasmid nicht hat, dem Rezipienten.
Nach der Kontaktaufnahme wird der Pilus abgebaut, bis sich die beiden Zellen berühren.
An der Berührungsstelle wird eine Konjugationsbrücke gebildet, deren Aufbau bisher nicht geklärt ist. Durch diese wird einer der beiden DNA-Stränge des Plasmiden in den Rezipienten übertragen.
Dies erfolgt bei ringförmigen Plasmiden meist über den rolling circle-Mechanismus, dabei wird ein DNA-Strang an einer spezifischen Stelle gebrochen, „abgerollt“ und in den Rezipienten transferiert. Bereits während des Abrollens wird der abgegebene Strang im Donor durch Synthese ersetzt. Nach der Übertragung wird der einzelne Strang im Rezipienten zum Doppelstrang ergänzt und die Plasmidform eingenommen. Am Ende der Konjugation besitzen beide Zellen das konjugative Plasmid und sind damit beide Donoren.
Bei der Übertragung des Plasmidstranges können auch weitere Plasmide, auch nicht konjugative, transferiert werden. Diese benötigen jedoch sogenannte mob-Gene, um den Pilus ebenfalls zu nutzen. Dieses Phänomen wird als „Mobilisierbarkeit“ bezeichnet.
Grampositive Bakterien
Grampositive Bakterien können die Konjugation anstatt über Plasmide auch über konjugative Transposons vermitteln. Im Gegensatz zu den gramnegativen Bakterien bilden sie keine Sexpili aus.
Die potentiellen Rezipienten scheiden Pheromone aus, die Donoren dazu anregen, ein sog. Aggregationsprotein zu bilden, durch welches sich die beiden Partner aneinanderheften können. Dann kann DNA übertragen werden. Die Vorgänge der Konjugation sind bei den grampositiven noch weniger erforscht als bei gramnegativen Bakterien.
Archäen
Auch bei Archäen der Gattung Sulfolobus wurden konjugative Plasmide entdeckt. Der genaue Ablauf der Konjugation ist hier jedoch kaum bekannt. Da die genetische Ausstattung der Plasmide sich erheblich von denen bakterieller Plasmide zu unterscheiden scheint, wird angenommen, dass sich auch die Mechanismen der archäalen Konjugation deutlich von denen der bakteriellen Konjugation unterscheiden.
Episome
Hat ein Plasmid bestimmte Sequenzen in der DNA, Insertionssequenzen (IS), die homolog zu Sequenzen im Bakterienchromosom sind, kann das Plasmid in das Chromosom eingebaut werden.
Solche Plasmide werden als Episome bezeichnet. Zellen, die ein Episom im Chromosom lokalisiert haben, werden hfr (hohe Frequenz der Rekombinationen) genannt. Da die Plasmidgene weiterhin vorhanden sind, können auch Konjugationen stattfinden, allerdings werden nun neben einem Teil der Plasmidgene auch chromosomale Gene übertragen. Diese können im Rezipienten mit der eigenen DNA rekombiniert werden. Da meist nicht alle Plasmidgene übertragen werden, wird der Rezipient in diesem Fall der Konjugation nicht zum Donor.
Übertragung chromosomaler Gene ins Plasmid
Ins Chromosom integrierte Plasmide können dort auch wieder ausgeschnitten werden. Da oft mehrere homologe IS im Chromosom vorliegen, können mit dem Plasmid auch chromosomale Teile mitausgeschnitten und in das Plasmid integriert werden.
Dieses veränderte Plasmid überträgt bei der Konjugation auch die chromosomalen Elemente, dadurch entsteht bei der Konjugation innerhalb desselben Stammes eine teilweise Diploidie. Da die Rezipienten hier das ganze Plasmid erhalten, werden sie nach der Konjugation zu Donoren.
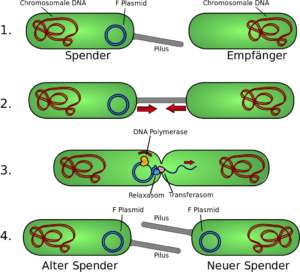
F-Plasmid
Meist wird die Konjugation am Beispiel des F-Plasmiden (F für Fertilität), das im gramnegativen Modellorganismus Escherichia coli vorkommt, erklärt. Dabei werden die Sexpili als F-Pili bezeichnet, der Donor wird F+ und der Rezipient F− genannt. Die Stelle auf dem Plasmid, an der der Transfer beginnt, ist die oriT-Sequenz. Ein mit chromosomalen Anteilen ausgeschnittenes F-Plasmid wird als F‘-Plasmid bezeichnet.
Gen-Kartierung
Mit der Zeitabhängigkeit der Übertragung der DNA eines hfr-Stammes kann man ermitteln, in welcher Reihenfolge sich die Gene auf dem Chromosom befinden. Dazu werden hfr-Stämme und Rezipienten mit bekannten Eigenschaften benutzt.
Die Übertragung der DNA auf den Rezipienten wird in verschiedenen Zeitabständen durch mechanische Kräfte gestoppt. Durch Minimalmedien, d. h. Nährböden auf denen sich nur Organismen mit bestimmten Eigenschaften vermehren können, kann dann festgestellt werden, welche Gene übertragen wurden. Die Zeitabstände ermöglichen die Einordnung, wo auf dem Chromosom sich das Gen befindet, deswegen sind ältere Genkarten oft in Minuten unterteilt.
Die Genkarte von Escherichia coli wurde lange vor der Sequenzierung durch diese Methode ermittelt.
Konjugation bei Ciliaten
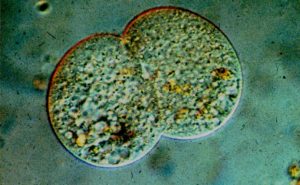
Als Konjugation bei Ciliaten bezeichnet man eine spezielle Form sexueller Vorgänge bei den einzelligen Wimpertierchen (Ciliophora). Dabei legen sich zwei Individuen aneinander und bilden eine sogenannte Plasmabrücke aus, durch die Genmaterial ausgetauscht wird.
Die Konjugation und der Austausch finden nur zwischen Individuen statt, die verschiedenen Paarungstypen angehören. So wird verhindert, dass Angehörige desselben Paarungstyps Genmaterial austauschen. Die Paarungstypen werden durch Glykoproteine auf der Oberfläche definiert.
Bei Ciliaten besteht das Genmaterial aus einem Mikronukleus und einem Makronukleus. Bei der Konjugation, löst sich der Makronukleus allmählich auf und aus den Mikronuklei beider Partner entstehen durch beide Teilungsvorgänge der Meiose jeweils vier haploide Kerne.
Bis auf jeweils einen dieser haploiden Kerne lösen sich alle so entstandenen Kerne ebenfalls wieder auf. Die beiden verbliebenen Kerne teilen sich nun in einer weiteren Mitose in zwei haploide Kerne, einen stationären Kern und einen Wanderkern.
Der stationäre Kern, auch als weiblicher Kern bezeichnet, bleibt im jeweiligen Individuum; der Wanderkern, auch als männlicher Kern bezeichnet, dringt über die Plasmabrücke in den Konjugationspartner ein und verschmilzt dort mit dessen stationären Kern. Damit hat nun jedes Individuum einen diploiden Kern.
Nach der Trennung beider Geschlechtspartner wird durch eine weitere Mitose der diploide Kern verdoppelt, aus einem der beiden Tochterkerne wird durch Polyploidisierung der Makronukleus aufgebaut, der andere Tochterkern bleibt unverändert als Mikronukleus.