Dictyostelium discoideum
- Wikipedie-de: Dictyostelium discoideum
Dictyostelium discoideum ist eine weltweit vorkommende Art der Schleimpilze und lebt als Einzeller im Boden. Ausgelöst durch Nahrungsmangel schließen sich viele der einzelligen Amöben zu einem vielzelligen Verband („soziale Amöbe“) zusammen und bilden an einem langen Stiel einen Fruchtkörper aus, aus dem Sporen entlassen werden.
Die Art ist seit ihrer Entdeckung im Jahr 1935 zu einem bedeutenden Modellorganismus der Biologie geworden.
Wachstumsphase
Dictyostelium discoideum ist in der vegetativen Phase eine haploide, einzellige und einkernige Amöbe von unregelmäßiger Gestalt und einem Durchmesser von rund 10 bis 20 Mikrometer. Eine Zellwand fehlt, die Zelle wird nur durch eine Zellmembran abgeschlossen. Sie weist zahlreiche Scheinfüßchen und Nahrungsvakuolen auf.
Bei ausreichender Verfügbarkeit von Nahrung lebt Dictyostelium discoideum als Einzeller und vermehrt sich durch Zellteilung. Es ernährt sich von Bodenbakterien, die es aufnimmt, indem es sie umfließt und so in eine Vakuole einschließt (Phagozytose).
Danach verbinden sich Vakuolen mit Verdauungsenzymen, den Lysosomen, und das Bakterium wird verdaut. Zwischen zwei Zellteilungen phagozytiert D. discoideum circa 1000 Bakterien.
Dictyostelium bewegt sich, indem es Zellfortsätze, sogenannte Scheinfüßchen bildet, die es nach vorne ausstreckt und dann den Zellkörper nachzieht. Wie bei den Bewegungen der menschlichen Muskelzellen funktioniert das auch hier durch die Zusammenarbeit von Aktin– und Myosin-Filamenten.
Mit diesen Bewegungen reagieren sie auf chemische Reize (Chemotaxis), also sozusagen auf den Geruch der Bakterien, von denen sie sich ernähren oder später bei der Schleimpilzbildung auf die Botenstoffe ihrer eigenen Artgenossen.
Hungerphase
Unterschreitet das Verhältnis zwischen der Menge an verfügbarer Nahrung und der Populationsdichte der Amöben einen kritischen Wert, geht D. discoideum aus der vegetativen Wachstumsphase in eine Entwicklungsphase über, die durch grundlegende morphologische Veränderungen und unterschiedlicher Genexpression gekennzeichnet ist.
Die Amöben sind in der Lage mittels eines Glykoproteins, dem Pre-Starvation-Faktor (PSF), das Verhältnis von Populationsdichte zu bakterieller Nahrungsquelle zu erkennen.
Dieser Faktor wird während der G2-Phase des Zellzyklus synthetisiert, sezerniert und akkumuliert im nahen Umgebungsfeld der Amöbe. Wird ein definiertes Konzentrations-Verhältnis überschritten, wird die Hungerphase eingeleitet, die den Transit zur Entwicklungsphase darstellt.
In der Hungerphase beginnen erst einige Amöben cAMP als Hungersignal auszuscheiden (sezernieren). Es wird unmittelbar über eine Signalkaskade die Entwicklungsphase eingeleitet. Der unmittelbare weitere Verlauf der Signalkaskade ist noch nicht genau aufgeklärt.
Entwicklungsphase

Streaming
Von einigen Amöben sezerniertes cAMP lockt andere Amöben an, die dann ihrerseits cAMP produzieren und sezernieren. Das sezernierte cAMP leitet die Chemotaxis ein, die dazu führt, dass die Amöben beginnen, sich in die Richtung der ansteigenden cAMP-Konzentration zu bewegen. Dies führt zur Ausbildung der typischen verästelten Bahnen. Aufgrund der cAMP-Ausschüttung auf der der Bewegungsrichtung abgewandten Seite der Zelle bildet sich eine „Head to tail“-Formation.
Pseudoplasmodium
In Folge aggregieren 50.000 bis 100.000 Amöben und formieren sich zu einem Pseudoplasmodium, auch „Mound“ genannt (engl. für „Hügel“). Dabei fusionieren die Zellen jedoch nicht, sondern bilden einen multizellulären Verband, der eine Populationsdichte von mindestens 400 Zellen/mm² hat. Bereits zu diesem Zeitpunkt kommt es zu einer Veränderung der Aktivität der entwicklungs-relevanten Gene und damit zu einer Differenzierung der Zellen in die zwei verschiedenen Zelltypen: Prä-Sporenzellen und Prä-Stielzellen.
Slug
Im weiteren Verlauf der Entwicklung entsteht aus dem „Mound“ ein „Slug“ (engl. für „Nacktschnecke“). Dieser Slug, von einer Schleimschicht umgeben, ist in der Lage, auf phototaktische, chemotaktische oder thermotaktische Reize mit Bewegung zu reagieren. Im Slug-Stadium erfolgt die Einstellung des Gleichgewichts aus circa 20 % Prä-Stielzellen und 80 % Prä-Sporenzellen und deren Sortierung. Die Sortierung wird über die unterschiedliche Sensitivität der zwei Zelltypen gegenüber cAMP gesteuert. Die Prä-Stielzellen sammeln sich aufgrund einer erhöhten cAMP-Sensitivität im vorderen Bereich des Slugs, während die Prä-Sporenzellen den restlichen Slug bilden.
Finger-Stadium
Im weiteren Verlauf kann der Slug gleich in die Kulmination übergehen oder aber umherwandern. Dies dient der Suche einer geeigneteren Stelle für die Kulmination. Dabei streckt sich der Slug in die Höhe (Finger-Stadium), um dann schneckenartig umherzuwandern (wandernder „slug“).
Kulmination
Im Lauf der weiteren Entwicklung wird an der Spitze des Slugs ein cAMP-Gradient ausgelöst, indem die Prä-Stielzellen vermehrt extrazelluläre cAMP-Phosphodiesterasen exprimieren. Die Abnahme der cAMP-Konzentration an der Slug-Spitze löst die Kulmination aus.
Mexican Hat
Es bildet sich an der Basis eine Stielröhre, an deren Kopfende sich die Prä-Stielzellen befinden („mexican hat“). Die Prä-Stielzellen schwellen durch Vakuolisierung an und sterben dann ab; gleichzeitig werden die Prä-Sporenzellen in die Höhe gehoben. Sie kondensieren durch Abgabe von Wasser, umgeben sich zum Schutz vor Hitze und Trockenheit mit einer Mukopolysaccharidhülle und gehen dann in einen Ruhezustand über. Der Sporenkopf wird durch die „upper“ und „lower cups“ am Stiel des Fruchtkörpers fixiert.
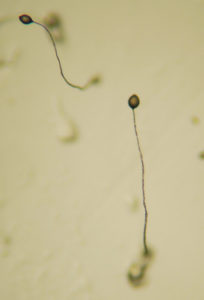
Fruchtkörper
Den Abschluss des Entwicklungszyklus bildet der Sporokarp („Fruchtkörper“). Dieser besteht aus toten Stielzellen, keimungsfähigen Sporenzellen und einer Basalscheibe zur Verankerung auf dem Substrat. Die ovalen Sporen sind gegenüber Hitze und Austrocknung unempfindlich und in der Lage, unter geeigneten Umweltbedingungen wieder als Einzeller auszukeimen und einen neuen Entwicklungszyklus einzuleiten.