Raues ER Das raue ER (rER), auch granuläres ER oder Ergastoplasma genannt, hat zwei Funktionen: die Proteinbiosynthese und die Membranproduktion. Seinen Namen hat es von den Ribosomen, die auf seinen Membranoberflächen sitzen. Es findet sich vorwiegend in den Zellen exokriner Drüsen und der Leber sowie in Nerven- (Nissl-Schollen) und Embryonalzellen.
- Wikipedia-de: Raues ER
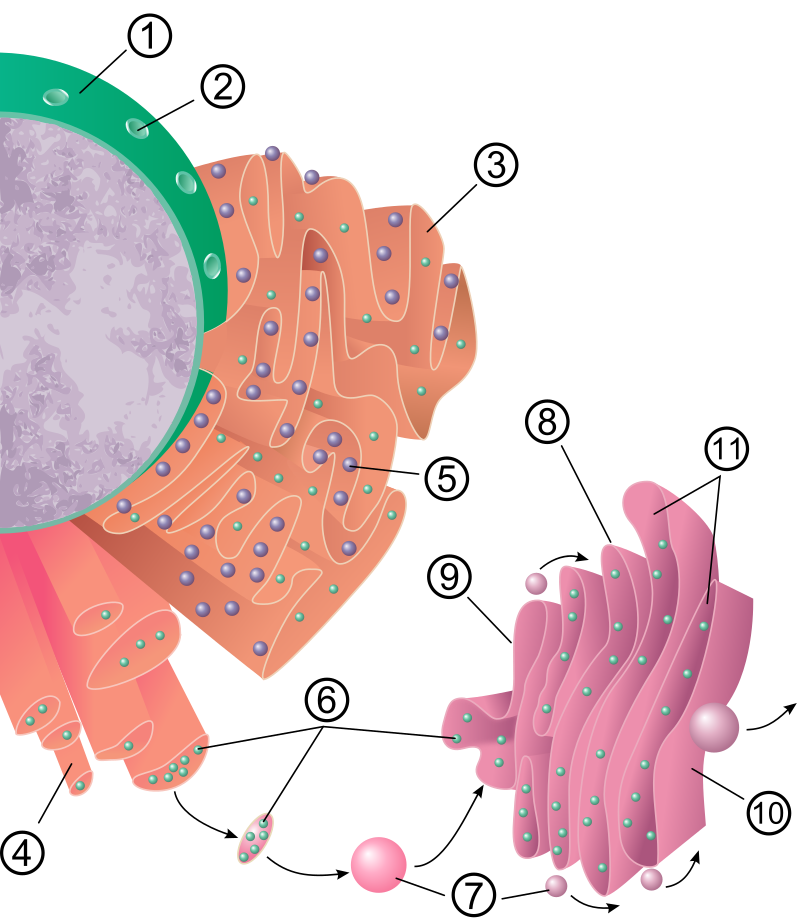
Das raue ER (rER), auch granuläres ER oder Ergastoplasma genannt, hat zwei Funktionen: die Proteinbiosynthese und die Membranproduktion.
Seinen Namen hat es von den Ribosomen, die auf seinen Membranoberflächen sitzen. Es findet sich vorwiegend in den Zellen exokriner Drüsen und der Leber sowie in Nerven- (Nissl-Schollen) und Embryonalzellen.
Das raue ER lässt sich mit basischen Farbstoffen wie Hämatoxylin, Kresylviolett oder Toluidinblau sichtbar machen (Nissl-Färbung).
Proteinbiosynthese
Proteine werden häufig von spezialisierten Zellen ausgeschieden (Sekretion). Diese Proteine werden von den Ribosomen produziert, die dem rauen ER anhaften. Eines dieser Proteine ist zum Beispiel das Insulin aus Zellen der Bauchspeicheldrüse.
Alle in membranengebundenen Ribosomen entstehenden Polypeptidketten werden zunächst in das Lumen des ER geschleust.
Dies geschieht durch porenbildende Proteine (Kotranslation). Auch im Zytosol synthetisierte Proteine werden in das Lumen des ER befördert (Posttranslation). Im Lumen des ER werden die Polypeptidketten zurechtgeschnitten und gefaltet.
Die linearen Aminosäureketten werden nach der Translokation in das ER gefaltet, erhalten also ihre dreidimensionale Struktur. Dieser Prozess wird von anderen Proteinen im ER unterstützt (Chaperone) und kontrolliert.
Fehlgefaltete Proteine werden umgehend retranslokiert, das heißt zurück ins Zytosol transportiert und dort durch das Proteasom degradiert. Das Cholera-Bakterium nutzt diesen Mechanismus, um sein Toxin über diesen Prozess in das Zytosol zu bringen, wo es aber der Degradation durch das Proteasom entkommt und seine toxische Wirkung entfalten kann.
Die meisten Sekretionsproteine sind Glycoproteine, welche kovalent gebundene Kohlenhydrate tragen. Diese Kohlenhydrate, es handelt sich um Oligosaccharide, werden im Lumen des ER durch die Enzyme des ER angeheftet. Die fertigen sekretorischen Proteine verbleiben im Lumen des ER und werden somit von Proteinen im Zytosol, welche von freien Ribosomen erstellt wurden, ferngehalten. Die sekretorischen Proteine werden in Form kleiner Membranbläschen abgeschnürt und verlassen so das Lumen des ER als Transportvesikel in Richtung Golgi-Apparat.
1999 erhielt Günter Blobel den Nobelpreis für Physiologie oder Medizin für seine 1975 gemachte Entdeckung, dass Proteine durch endogene Protein-Signale (Signalsequenzen) vom ER aus in verschiedene Zellkompartimente zielgerichtet weitergeleitet werden.
Als Signalsequenz in diesem Sinne wird eine bestimmte N-terminale Peptidsequenz bezeichnet, die nach dem Transport durch die Membran des ER durch die Signalpeptidase abgespalten wird.
Proteine, die für Ziele außerhalb des ERs bestimmt sind, werden anschließend in Transportvesikel verpackt und entlang des Zytoskeletts zu ihrem Bestimmungsort weitergeleitet.



Impressum